Interesanti par putniem....
http://tencinu.wordpress.com/2012/07/04 ... korotkovs/
...
Zinātnieki pārsteigti uzzināja, ka galveno enerģiju putni tērē nevis lidošanai, bet ēšanai.
...
Atklāts, ka pārlidojumā strazdi gandrīz nezaudē svaru (zudumi ir mazāki par 6% no svara), bet tauku daudzums paliek nemainīgs. Gaidītais, ka tie enerģiju iegūst, tērējot savus taukus, nepiepildījās. Putniņi tiešām pirms lidojuma palielina tauku daudzumu, bet tie kalpo galvenokārt siltuma saglabāšanai. Jo augstāk, jo aukstāks, un lai netērētu papildus enerģiju siltumam, putni veido “siltumizolāciju”, mētelīti, kas tos lidojumā sargā no aukstuma.
Veiksim vienkāršus aprēķinus. 30 gramus smagam putnam 6% ir 1,8g. Sadedzinot 1g ogļhidrātu, izdalās 4 kilokalorijas enerģijas (16,75 kDž), 1g tauku dod 9kkal jeb 37,68kDž. 1,8g ķermeņa masas tieša pārvēršana enerģijā var nodrošināt no 30 līdz 68kDž. Īstenībā šis skaitlis ir daudz mazāks, jo pārvēršanas koeficients organismā ir daudz mazāks par 100%.
Kā redzam, no 130kDž, kas vajadzīgi lidojumam, ar barības krājumu sadedzināšanu var nodrošināt mazāk par pusi.
Jo vairāk tas attiecas uz putniem, kas lido pār Atlantijas okeānu. Piemēram, sarkanspārnu tārtiņi, kuru spārnu vidējais vēziens ir vien 34cm, savās sezonas migrācijās veic beznosēšanās 3300km pārlidojumu ar ātrumu ap 90km/st no Aleutu līdz Havaju salām. No klasisku priekšstatu viedokļa tāds lidojums nav iespējams, putniem būtu jānobeidzas no novājēšanas kaut kur pusceļā.
No klasisku priekšstatu viedokļa tāds lidojums nav iespējams, putniem būtu jānobeidzas no novājēšanas kaut kur pusceļā.
Tātad, no klasiskā redzes viedokļa putnu migrācijas lidojumi nav iespējami!
Tomēr putni par to neko nezin un pārlido jau tūkstošiem gadu.
Vai tiešām putni darbojas pēc kādiem citiem, mums nezināmiem fizikas likumiem?
Mēs ļoti piesardzīgi izturamies pret “īpašu bioloģisku likumu jeb īpašas bioloģiskas enerģijas” apspriešanu. Jebkuru neizprotamu norisi vispirms jāmēģina izskaidrot ar zināmām likumsakarībām. Mūsdienu bioloģijā ļoti lielas izredzes ir SINERĢĒTIKAS – zinātnes par atvērtām termodinamiskām sistēmām – likumsakarību lietojumam. 19.gadsimta klasiskā zinātne visas sistēmas aplūkoja kā SLĒGTAS – tādas, kas pastāv iekšējās norisēs. Šāda pieeja noveda pie daudziem strupceļiem termodinamikā un fizikā. Pretrunas atrisinājās, pateicoties Iļjas Prigožina un Hakena zinātnisko skolu darbiem. Viņi pierādīja, ka vairums īstenu dabas sistēmu ir jāuzskata par ATVĒRTĀM – tādām, kas pastāv apmaiņā ar apkārtējo telpu – ne vien ar vielu, bet arī ar enerģiju un informāciju. Pie kam ar pēdējiem katrā noteiktā gadījumā var saistīt noteiktas fizikālas norises. Aplūkosim enerģijas izguves mehānismu no gaisa, piemēram, kā tas ir putniem.
Tā ir arī ar bioenerģiju. Daudz jau zinām, daudz saprotam, bet tā nav pēdējā patiesība. Un izziņas ceļā mūs vēl gaida daudz brīnumainu atklājumu. Bet atgriezīsimies pie putniem.
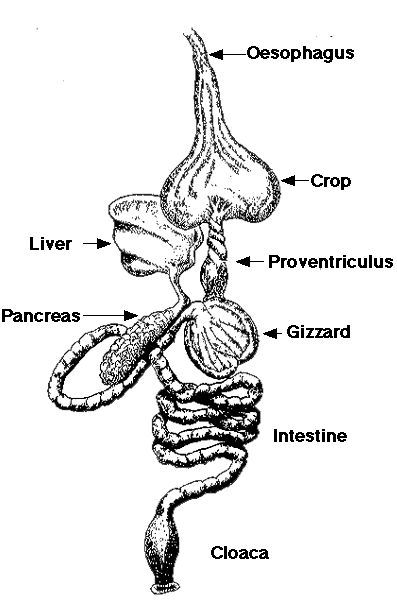
Putnu elpošanas sistēma ir ja ne vispilnīgākā, tad vissarežģītākā no visiem mugurkaulniekiem. Elpvados neizmantots gaiss ir vien trahejā, bet caur plaušām gaiss plūst tikai vienā virzienā, pie kam pilnu ciklu gaiss iziet divos elpošanas pāros (ieelpa-izelpa, ieelpa-izelpa), tā sauktajā dubultelpošanā. Bronhas, ieejot plaušās, nodod gaisu otrās pakāpes bronhās, kas daļēji iziet ārpus plaušām un veido gaisa maisus, kas izvietoti dažās putna ķermeņa vietās. Otrās pakāpes bronhas savā starpā savieno daudzas parabronhas, kas apvītas ar asinsvadu kaplāru tīklu. Gaisa maisu tilpums ir vairākas reizes lielāks par plaušu tilpumu. Gaisa maisi ir izvietoti starp iekšējiem orgāniem, starp muskuļiem, zem ādas, un ir savienoti ar vairākiem kaulu dobumiem. Gāzu apmaiņā gaisa maisi nepiedalās, tie pilda citus uzdevumus, tostarp nozīmīgākie ir plaušu ventilācija un siltumatdeve. Galvenā putnu elpošanas īpatnība – plaušas, kas netiek izplestas, ir slēgtas cietā krūšu kurvī, kas nemaina tilpumu. Tāpēc plaušas tiek gaisa caurpūstas ar bronhu sistēmas palīdzību, bet gaisa plūsmu uztur elpmaisu tilpuma izmaiņa.
Zīmīgi, ka putnu elpvados nav atklāti nekādi vārsti, un visa savādā gaisa kustība notiek saskaņā ar hidrodinamikas likumiem. Gāzu apmaiņas straujumu veicina asinsrites pretplūsmas sistēma putnu plaušās, tas ir, asinis un gaiss plūst pretējos virzienos. Tādēļ “svaigāks” gaiss saskaras ar “arteriālākām” asinīm, kas nodrošina efektīvu gāzu apmaiņu. No 1 litra gaisa putni izgūst 40 mililitrus skābekļa (zīdītāji — 30ml), pie tam skābekļa spraigums (напряженность) arteriālajās asinīs ir lielāks, bet ogļskābās gāzes spraigums mazāks kā izelpojamā gaisā!
Tālākās šīs nodaļas lappuses līdz nodaļas secinājumiem cilvēkiem, kuri ne visai interesējušies par bioloģiju, var izrādīties pārāk apgrūtinošas, tāpēc tās var bez liela zaudējuma izlaist. Galvenā doma ir, ka mūsdienu zinātniskie priekšstati ir pareizi, bet ne vienmēr pilnīgi pabeigti. Zinātne nav sastingusi pilnīga sistēma, bet dzīvs organisms, kas pastāvīgi attīstās un sasniedz jaunus apvāršņus. Idejas ar vislielākajām izredzēm dažkārt laikabiedriem šķiet neprātīgas, bet vēlāk pie tām pierod un uztver bez brīnīšanās.
Skābekļa molekulai piemīt daudz unikālu īpašību. Tai ir divi elektroni ar paralēliem spiniem valences molekulārajā orbitālē (М ^^, kur zīme ^ nozīmē elektronu ar noteiktu spina virzienu). Tāds ārējā elektronu apvalka stāvoklis tiek saukts par tripletu. Tripleta skābeklis ( 3O2 ; tas ir skābekļa molekulas pamatstāvoklis) ir potenciāls enerģijas avots, tomēr tā nevar īstenoties patvaļīgi, jo atbilstoši Vīgnera saglabāšanās likumam tieša reakcija ar molekulām singletā (singlets skābeklis jeb 1O2 ir molekulāra skābekļa elektroniski ierosināts stāvoklis, kas ir nenoturīgāks kā parastajam tripleta skābeklim) stāvoklī nav iespējama. Tas ir viens no tripleta skābekļa noturīguma nosacījumiem. Ir vairāki skābekļa aktivizācijas veidi, un viens no tiem ir vienelektrona vairošanās. Skābeklim pārtverot elektronus, rodas starpprodukti – skābekļa aktīvās formas (ROS, reactive oxygen species). Daļa no tām ir brīvie radikāļi – molekulas ar elektronu nepāra skaitu ārējās orbitālēs. Lai iegūtu otru elektronu, tādas molekulas aktīvi mijiedarbojas ar apkārtējām molekulām, kas tad kalpo par elektronu donoriem. Brīvais radikālis satver elektronu un pārvēršas par molekulu, kamēr tā donors sāk meklēt savu donoru. Tādā veidā brīvie radikāļi var izraisīt ķēdes reakciju tādos organisku molekulu kā lipīdi, proteīni, ogļhidrāti šķīdumos. Optimāla vide tādām norisēm ir asinis.
Radikāļu ķēdes reakcijas tiešām var kaitēt bioloģiskām molekulām mēģenē (in vitro), tāpēc skābekļa aktīvās formas bioloģiskajā un medicīniskajā literatūrā tiek uzskatītas par veselībai kaitīgu faktoru. Tomēr liels datu apjoms liek domāt, ka īstenībā skābekļa aktīvās formas ir bioloģiskās norises nozīmīga sastāvdaļa. Saskaņā ar dažādiem vērtējumiem 10-15% patērētā skābekļa miera stāvoklī tiek pārveidoti vienelektrona mehānisma norisē, kā gaitā rodas skābekļa aktīvās formas. Spriedzes vai aktīva darba apstākļos, kad skābekļa aktīvās formas ražojošo enzīmu aktivitāte palielinās, skābekļa patēriņš palielinās par 20-40%, un viss šis virsnormas daudzums tiek pārveidots vienelektrona mehānisma norisē. Tātad skābekļa aktīvajām formām vajadzētu būt nozīmīgai lomai normālā fizioloģijā.
Būtiski svarīgi ir, ka rekombinācijas norisē izdalās enerģijas kvanti, kas ekvivalenti redzamā vai ultravioletā diapazona enerģijai. Galvenais donors šajās norisēs ir ūdens molekulas, kurām ir vismazākais jonizācijas potenciāls. A.Gurvičs un E.Sent-Djerdji neatkarīgi viens no otra uzrādīja, ka dzīvās sistēmās elektronu ierosinājuma enerģija neizkliedējas siltumā, bet tiek nodota ar makromolekulām vai to ansambļiem. Enerģijas porcijas, kas ekvivalentas fotonu enerģijai, kas rodas radikāļu rekombinācijas reakcijās, var kalpot par bioķīmisko reakciju palaidējiem (trigeriem), nodrošinot metabolisku norišu ritmisku raksturu.
Katrā ķēdes reakcijā atbrīvojas līdz 8 elektronvolti (EV) enerģijas. Putnam, kas lidojot ieelpo miklu gaisu, tas rada papildus enerģijas avotu adenozīntrifosfātu ražošanai muskuļos. 1EV ir vienāds ar 1.6х10-19 džouliem, vienā gaisa kubikcentimetrā ir apmēram 1016 molekulu, tāpēc 100kDž enerģijas ražošanai putnam ir jāpārstrādā apmēram 106 kubikcentimetri gaisa. Putna elpošanas biežums lidojumā ir 60-160 cikli minūtē. Tāpēc šā gaisa apjoma caurlaišanai putnam vajag 102-103 minūtes jeb 2 līdz 16 stundas. Šis laiks ir salīdzināms ar migrācijas lidojuma ilgumu.
Aprakstītais mehānisms nepretendē uz pilnību sīkumos, bet tas ļauj izdarīt dažus secinājumus:
. Mūsdienu biofizikas koncepcijas vēl tikai veidojas, aina ir tālu no pilnības.
. Bioloģiskie organismi izgūst enerģiju ne vien no barības, bet arī tieši no gaisa, ūdens un gaismas.
. Ārējiem stimuliem ir pamatliekoša loma dzīvības iekšējo norišu aktivācijā. Citiem vārdiem, supervāji informatīvi stimuli aktivizē homokinētisku ķēdes reakciju kaskādes.
Izklāstītie pamatlikumi kaut kādā mērā ir lietojami arī cilvēka organisma energoapgādes norisēm, īpaši aktīvas darbības, piemēram, ilgstošu sportisku sacensību gaitā. Piemēram, triatlona sacensībās sportisti iztērē milzīgu enerģijas daudzumu bez iespējas to būtiski papildināt. Dabiski, aplūkotais mehānisms prasa sīku izpēti attiecībā uz cilvēka organismu, tomēr šādā pieņēmumu variantā tas ļauj iezīmēt praktiskus sportista organisma energoapgādes palielināšanas veidus.